

IT has been known for a long time that plants have electrodynamic properties. Their ability to process complex test currents and to behave in a computer-like "go/no-go" binary mode is unique. But never in all those millennia since the first green leaves poked their heads out of Paleozoic swamps have plants been given more professional attention (other than from botanists) than they have in recent years. After much initial skepticism regarding plants semi-conductive and general electromotive qualities, here are but a few of the topics that science is speculating about today:
1. Can plants be integrated with electronic readouts to form major data sensors and transducers?
2. Can plants be trained to respond to the presence of selected objects and images?
3. Is their alleged supersensory perception (SSP) verifiable?
4. Of the 350,000 plant species known to science, which family is most promising from an electronics point of view?
Electrical Characteristics
The effect of electrical excitation on plants was noted in the 18th century and described in Dr. Jallabert's book "De l'électricité des végétaux," published in Geneva in 1748. Later, in 1924, Dr. Stern gave an excellent summary in his publication "Die Elektrophysiologie der Pflanzen." Up-to-date findings were described in G. Ungar's work entitled "Excitation" (1963) which also gives a good overview of bioelectric and electrophysical phenomena in general. However, taken together, many schools continue to attach more significance to plant growth hormones and regulators than to electrical phenomena per se.
The behavior of a living cell is dramatically complex and unique. There are responses to light, heat and cold, radiation, injuries, and touch. Electrical properties can be investigated with microelectrodes, a sample of which is shown in the test arrangement of Fig. 1.
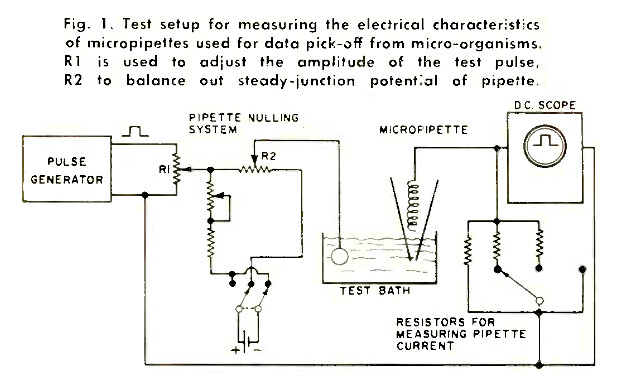
Microelectrodes usually take the form of micropipettes, consisting of a very thin glass envelope, a conductive liquid and a metallic helix or insert for conveying current to external readouts. Their electrical impedance is of the high ohm type and must be verified frequently by using a special test bath. The d.c. nulling arrangements and parallel resistances permit investigations of over-all impedance characteristics, which, ideally, should lead to no impairment of pulsing test currents propagated through the electrode systems. The test bath may be used as an approximate standard, featuring impedance and conductive properties similar to those of biological electrolytes.
Typically, strong electric currents flowing through a plant cell have the effect of causing an immediate contraction of the cytoplasm of a cell (best seen in the spirogyra or clodea catladcnsis), from which physiological recovery is impossible. Less intense currents cause either a partial retraction from the cell wall or, after the current has been stopped, resumption of normal cell functions. Upon swarm spores and antherozooids swimming in water, electricity has the peculiar effect of polarizing them. The one or the other extremity might turn in a definite direction, either with or against the direction of d.c. potentials.
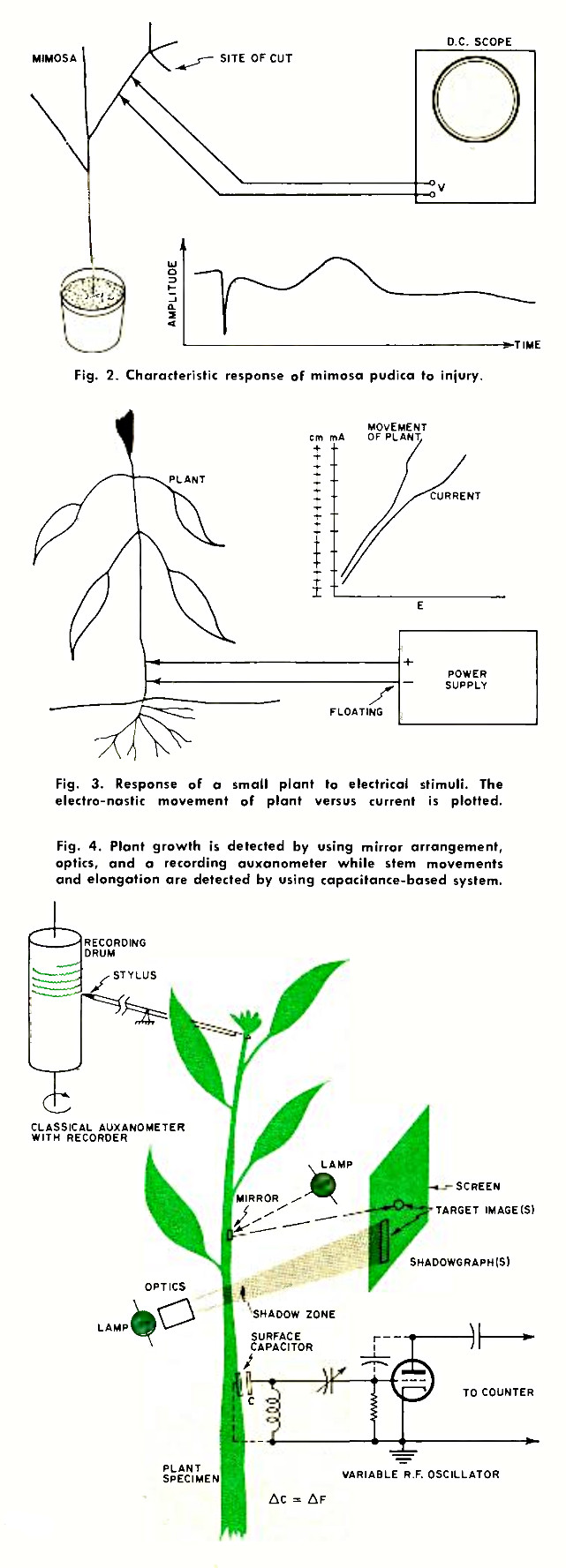
However, a different situation arises when sensitive plants, such as the mimosa pudica, experience injury. Fig. 2 shows an experimental setup used for electro-culture work, consisting of a high gain d.c. oscilloscope as data sensor. If the plant is cut, as shown, a typical current spike will ensue which is indicative of a nastic response. The term electro-nastic movement is used if, as in Fig. 3, application of current elicits a motive response. In the case of the mimosa, there exists no proportionality between the strength of stimuli and the dimensions of motive reaction. Thus, if a certain current threshold is exceeded, we will notice a total, "all-or-nothing" reaction. The specimen acts like a silicon controlled rectifier in that certain triggering levels must be present to cause action.
Electro-nastic performance is best seen on small plants. For this purpose, various plant-response indicators have evolved, working on different detection principles.
As shown in Fig. 4, simple growth can be detected with a recording auxanometer. Mirror arrangements and optics may be employed to verify movements by reflection of light and shadowgraphic methods.
The capacitance-based system is suitable for measuring stem movements and elongation. However, a device such as this is critical in that strong oscillator currents and/or d.c. must not act as a stimulus to the plant. This requirement can be met by carefully grounding zero-potential electrodes in the plant's pot and by adequate distance between acting capacitor members. Experimenters normally work with a "center frequency" of given value. If the center frequency is, say, 20 MHz and finally changes to 20.80 MHz while the experiment is in progress, the net value will be 800 kHz. By measuring the distance between capacitor plates (C) with a micrometer, it is possible to plot distance versus frequency as a result of growth or movement of the specimen.

Packaged Plant Electronics
The majority of electrical plant reactions occur in the millivolt range and involve long time bases. In electro-culture experiments with bean seedlings, for example, waveforms emanated by roots (Fig. 5) are complex and require careful shielding from industrial interferences. Since it is very difficult to properly "terminate" biological impedance, like a TV antenna, special packaging concepts and designs have emerged to keep plant-generated electrical data as "clean" as possible.
Fig. 6 shows an example of a special packaging concept. The living plant is housed in a quasi-Faraday cage to permit access of light and air. Light is important since the photo-reactions of the specimen normally require electromagnetic energy between a wavelength of 2000 and 10,000 angstroms (A). However, this particular experimental setup is unique in that it permits simultaneous biochemical and bioelectric observations of a plant.
In the case at hand, weak electrical signals are generated by shielded electronics in a special enclosure. These products consist of r.f. below 10 meters and pulsed d.c. Galvanic feedback loops permit immediate stoppage of currents if excitation levels are exceeded. Timing is provided by motor-driven switches beneath the cage, with current normally applied during daylight hours only.
Interest in the plant's chemical behavior is coupled with the above, but is more complex. One of them is the observation of auxins, which are chemicals (plant hormones) which cause distinct growth patterns. One of these substances, in-dole-3-acetic acid, occurs in minute quantities in growing plant tissue. Thus, in a shoot of the pineapple plant, only 6 micrograms of auxin are found per kilogram of plant material which is comparable to the weight of a needle in a 22-ton truckload of hay. There are other plant hormones and regulators, including gibberellins and cytokinins, that contribute to the electrically stimulated plant's well-being and enable it to deal with a massive set of physical forces. These aspects are of immense complexity and cannot be nailed down in laws and predictive formulas. There is much randomness involved here, which frequently makes a plant's behavorial response just as unpredictable as that of man. However, it is possible to come up with some guidance in the form of statistics and other laboratory data if the plant specimens have a reasonably controlled environment and are fed specific nutrients. A respective feeder can be seen at the upper right-hand side of the cage in Fig. 6.
The Backster Effect
Some ten years ago, in the "Moon Garden" developed by Republic Aviation at Farmingdale, N.Y., scientists were able to induce what appeared to be "nervous breakdowns" and "complete frustration" in plants being tested as possible space foods. At East Grinstead, Sussex, England, Dr. L. Ron Hubbard also noted electrogalvanic responses in tomatoes when struck by a nail. Unable to arrive at a reasonable explanation for these "findings" the investigators arbitrarily entitled these reactions "emotion-like" responses in plants.
Three years ago this "emotion-like" response in plants was noted by Clyde Backster, executive director of the Backster Research Foundation (N.Y.) and a polygraph (lie detector) expert, when he connected one of his instruments to a draena massangeana plant while it was taking up water. The specimen's psychogalvanic reflex (PGR) reaction pattern was strange, for it appeared to be similar to that of a human being undergoing emotional stimulation. Attempting to further test the plant's reaction, Backster decided to take a match and burn the leaf carrying the polygraph electrodes. At the moment this thought entered his mind a dramatic change was reflected in the PGR tracing - the graphic recorder suddenly drew a waveform of great amplitude. This totally unexpected reaction suggested to Backster that somehow the presence of this destructive mental image set off a chain of events, not obvious to the human senses, that triggered the plant into what seemed to be an emotion-like response. Later, in an attempt to substantiate his previous findings, Backster devised a complex experiment involving the dumping of live shrimp into boiling water. The experiment was executed at a random time by machines and, what was most significant, without Backster's presence. Again the plants selected for this experiment seemed to register what could be construed as a profound emotion-like response at the precise moment of the shrimps' death.
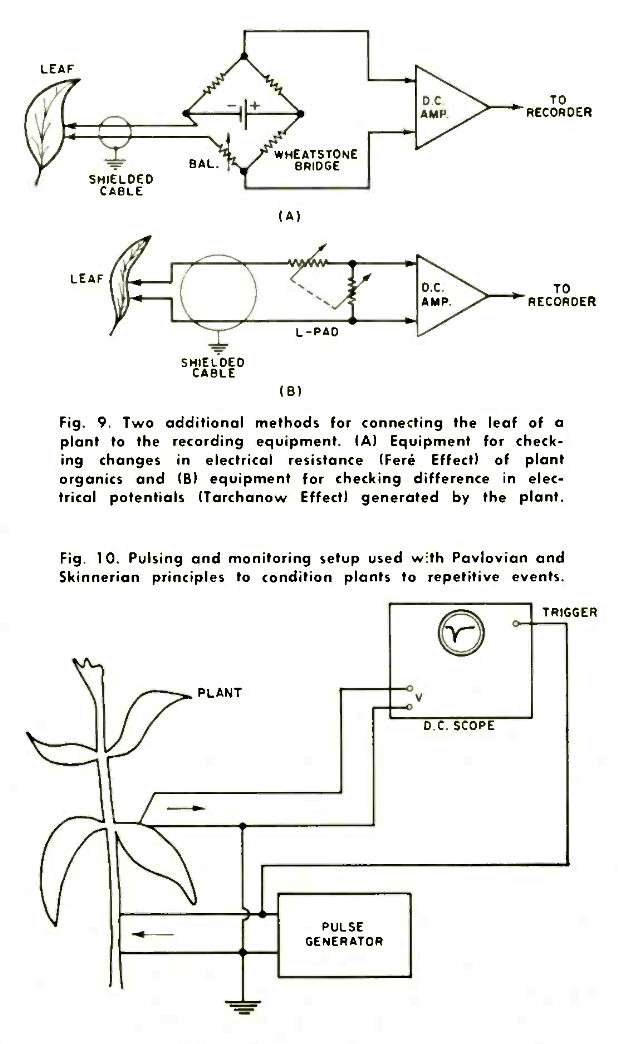
The basic equipment inventory required for verification of what is called the Backster Effect is shown in Fig. 7. The apparatus consists of the plant itself as the prime "data sensor," a differential d.c. preamplifier, and the graphic recorder. In a typical test situation, the amplifier is brought into close proximity to the plant (Fig. 8) and connected by shielded lines to the leaves. Fig. 9 shows two additional methods of connecting the leaf through the d.c. amplifier to the recorder. The recorder itself, set up remotely, requires little else but multiple-speed drive trains in order to obtain good trace resolution at various paper speeds.

However, it is only fair to point out that working with the Backster Effect involves much more than the mere ability to construct top-quality electronic equipment. There are certain qualities involved here which do not enter into normal experimental situations. According to those experimenting in this area, it is necessary to have a "green thumb" and (most important) a genuine love for plants.
Plant Conditioning
Experiments, such as the one shown in abstract form in Fig. 10, have been performed to demonstrate that plants can be conditioned to react to certain events. Those investigating this possibility feel that these life forms have memories and being living systems with many sensory capacities, are capable of being trained by Pavlovian and Skinnerian conditioning techniques. Unfortunately, unlike experimental situations involving dogs (Pavlov) and pigeons (Skinner), no specific reward systems - such as food - have as yet been worked out with plants; but experiments are under way.
Attention, of late, has also been given to larger plants as well. The yucca plant is a representative example of a plant too large to be examined indoors. The equipment shown in Fig. 11 was devised to test its responses. The instrumentation revolves around a high-gain d.c. amplifier and a d.c.-excited Wheatstone bridge. A galvanometer serves as an "emotion" readout, being calibrated in statements such as "Decrease" and "Increase." This article has given a fair sample of what has been and can be done in this new and exceptionally promising field. The almost human-like sensory and response characteristics attributed to plants by Clyde Backster and other investigators in the field are too unique to be ignored.
(Editor's Note: Although some of the items discussed may be controversial, we believe that our readers will find the story both interesting and stimulating. We would also like to mention that the Backster Effect, dealing with the electrical response of plants under "emotional" stress, was recently demonstrated on nationwide TV. The effect was also demonstrated "live" to one of our editors.)
REFERENCES
- Berry, L. J. & Hoyt, R.C. "Polarization and Stimulation of the Onion Root by Direct Current," Plant Physiol. 1943, 18:372-396. <http://plantphysiol.org/content/18/3/372.full.pdf>
- Clark, W.G. "Electrical Polarity and Auxin Transport." op cit. 1937. 12:409-410. <http://plantphysiol.org/content/12/3/737.full.pdf>
- Hollaender, A. (ed): "Radiation Biology, Vol. III", McGraw-Hill, 1956.
- Loehr, F.: "The Power of Prayer on Plants," New American Library. Signet Books, 1969.
- Lund. E.J.: "Bioelectric Fields and Growth," University of Texas Press, Austin, 1947.
- Machlis, L. & Torrey, J.G.: "Plants in Action - A Laboratory Manual of Plant Physiology," W.H. Freeman Co., San Francisco, 1956.
- McGraw, W.: "Plants are only Human." Argosy, pp. 24-27, June, 1969.
- Purvis, M.J. et al: "Laboratory Techniques in Botany," Butterworths. London. 1964.
- Sinyukhin, A.M. & Gorchakov, V.V.: "Role of the Vascular Bundles of the Stem in Long-Distance Transmission of Stimulation by means of Bioelectric Impulses," Soviet Plant Physiology, 15:3, pp. 477-487, May-June 1968.
- Suckling, D.: "Bioelectricity," McGraw-Hill, 1961.
- Thomas, J.B.: "Electrical Control of Polarity in Plants," Rec. Trav. botneerland, (Holland). 1939, 36:373-437.
- Ungar. G.: "Excitation," C.C. Thomas, Springfield. Ill. 1963.
- Went, F.W.: "The Experimental Control of Plant Growth," California Institute of Technology, 1957.
Originally published in "Electronics World" (Oct 1969). Browse AmericanRadioHistory.com for complete issues of E.W. and many other historical radio and electrical publications in PDF format.
Return to the BIO-ICOMM Project homepage or the Bibliography of L. George Lawrence.